deLemus
Dynamic Expedition of Leading Mutations in SARS-CoV-2 Spike Glycoprotein
Spike Glycoprotein
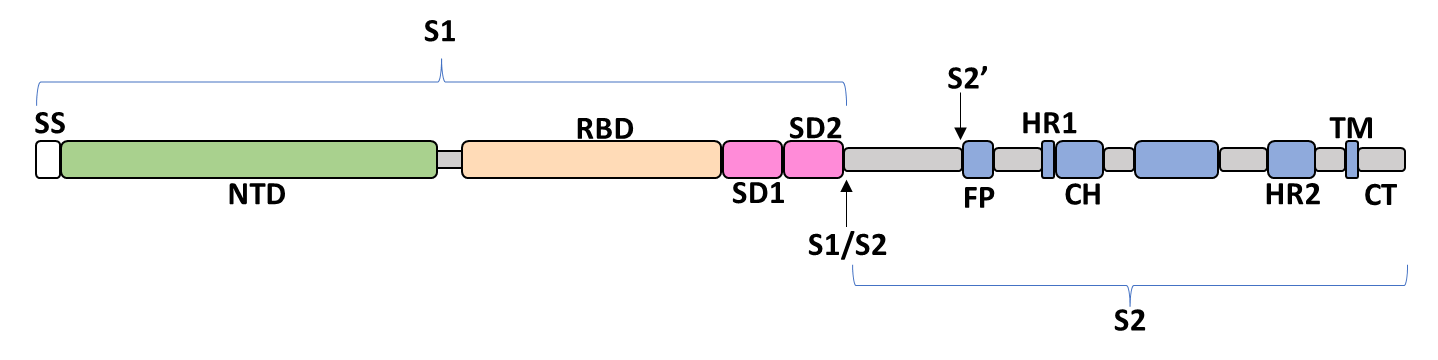
The spike glycoprotein of the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is a trimeric type I viral fusion protein that binds the virus to the angiotensin-converting enzyme 2 (ACE2) receptor of a host cell.[1] It is composed of 2 subunits: the N-terminal subunit 1 (S1) and C-terminal subunit 2 (S2), within which multiple domains lie. The S1 region facilitates ACE2 binding and is made up of an N-terminal domain (NTD ~ 1 – 325), a receptor-binding domain (RBD ~ 326 – 525), and 2 C-terminal subdomains (CTD1 and CTD2 ~ 526 – 688), while the downstream S2 region is responsible for mediating virus-host cell membrane fusion.
Update (03/02/2023)
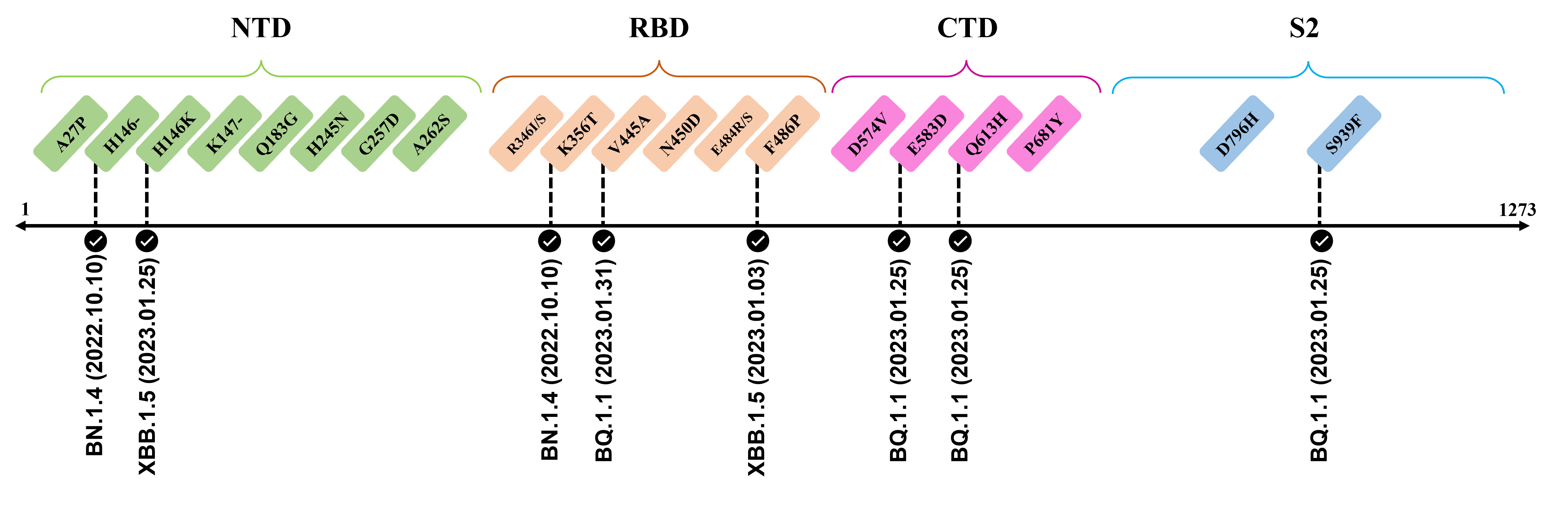
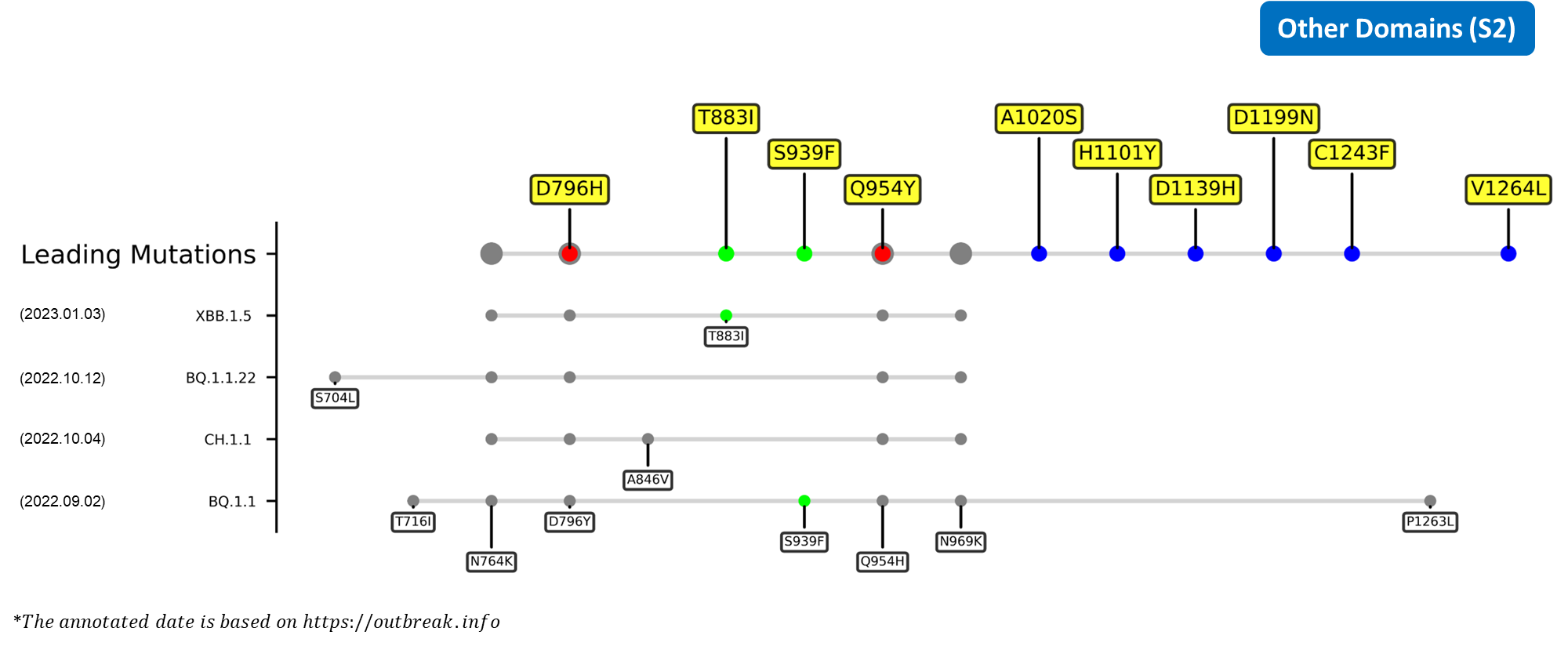
The recently confirmed leading mutations are listed as follows.
2023.01.31
| Mutation | Information |
|---|---|
| V445A | Confirmed in BQ.1.1 ; Amino acid site located at an RBD epitope[2] ; Mutation reduces neutralization by antibody [3] |
2023.01.17 - 2023.01.25
| Mutation | Information |
|---|---|
| H146-/K | Reported in BQ.1.1 and XBB.1.5 ; Amino acid site recognized by mAbs targeting NTD[4] |
| E583D | Shows up in BQ.1.1 ; Viral functions to be confirmed by further investigation |
| Q613H | Emerge in BQ.1.1 ; Speculate to enhance replicative fitness and transmissibility due to close proximity to D614G ; Potential functions to be elucidated[5][6] |
| S939F | Observed in BQ.1.1 ; Destabilize both pre-fusion and post-fusion S2 conformation[7] ; Capable to enhance infectivity and modulate T-cell immune response when combined with D614G[8][9] |
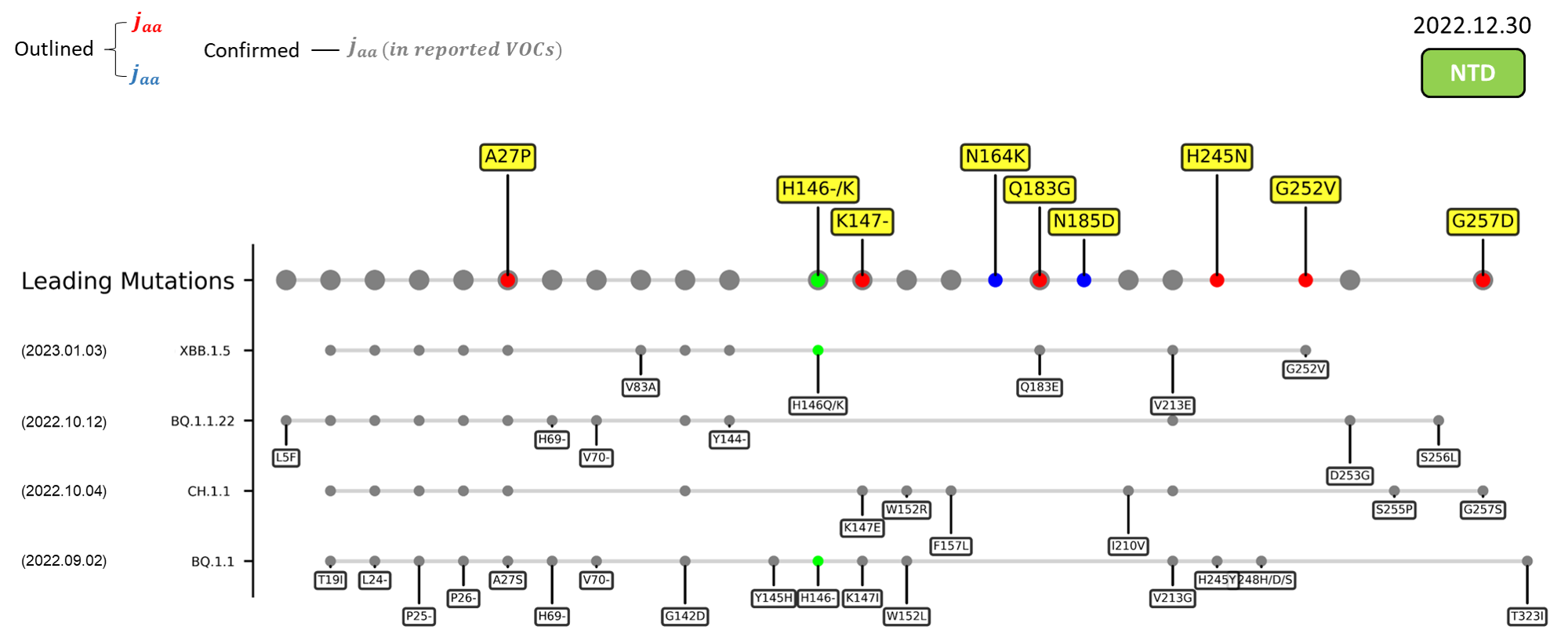
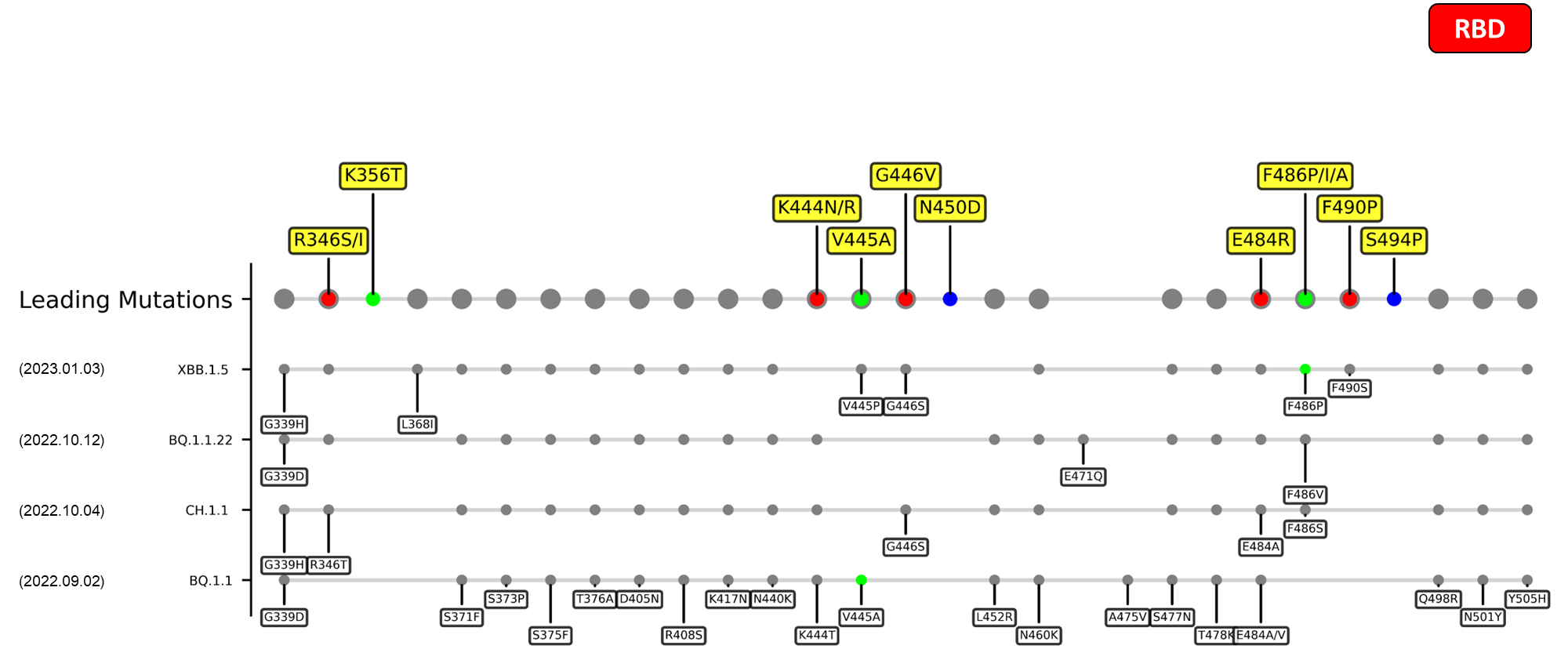
The following leading mutations call for special attention with respect to the upcoming variants.
NTD
| Mutation | Information |
|---|---|
| A27P | An antigenic site targeted by the group 3 antibody C1717[10] |
| K147- | Involved in interacting with multiple monoclonal antibodies[11] ; Mutation to threonine (K147T) at this site promotes immune evasion[4] |
| N164K | Functional impact to be confirmed in future investigation. |
| Q183G | Interactions with surface glycoconjugates mediate the viral attachment[12] ; Caused a loss of an amide group; May abrogate the hydrogen bond between the amino acid and the carboxylic group of surface sialosides[13] |
| N185D | Functional impact to be confirmed in future investigation. |
| H245N | Located in the supersite loop of the NTD antigenic supersite for antibodies SLS28 and S2X333[11][4] ; Caused a loss of a positive charge ; Introduces an NXS sequon (245NRS247) for N-glycosylation |
| G252V | Site is critical for the binding of human antibody COV2-3439[14] |
| G257D | Located in the supersite loop of the NTD antigenic supersite for antibodies SLS28 and S2X333[11][4] ; Caused a gain of negative charge |
| A262S | Enhance the utilization of ACE2 in numerous mammals[15] ; May increase interspecies and intraspecies transmissibility |
RBD
| Mutation | Information |
|---|---|
| R346I/S | Possibly lead to immune evasion due to the disruption of class 3 antibodies binding site[16] [17] |
| K444N/R | TBA. |
| G446V | TBA. |
| N450D | Results in antibody resistance[18] |
| E484R/S | A site of mutation being reported in multiple variants, mutation at this site could harbor escape mutations that impede the binding and neutralization ability of antibodies[5] [19] |
| F490P | Mutation at this site enable antibody escape over mAb COV2-2479, COV2-2050, COV2-2096 based on DMS study.[19] |
| S494P | This mutation persistently shows up in an immunocompromised patient of COVID-19, which was treated various drugs e.g. remdesivir, intravenous immunoglobulin, etc.[20] |
CTDs
| Mutation | Information |
|---|---|
| T547I | Functional impact to be confirmed in future investigation. |
| T572I | Functional impact to be confirmed in future investigation. |
| D574V | Located at the CTD1 region, substitution to an electrically neutral valine residue permits the endosomal entry efficiency and immune evasion ability of SARS-CoV-2.[21] |
| E619Q | Functional impact to be confirmed in future investigation. |
| E658S | Functional impact to be confirmed in future investigation. |
| I666V | Functional impact to be confirmed in future investigation. |
| S673G | Functional impact to be confirmed in future investigation. |
| P681Y | Located at the C-terminal of the CTD2, this substitution can diminish the cleavage efficiency of the S1/S2 interface because the bulky nature of tyrosine hinders the binding of furin to the cleavage loop.[22][23] |
| I688V | Functional impact to be confirmed in future investigation. |
S2
| Mutation | Information |
|---|---|
| D796H | Located in S2 region, the single aspartic acid-to-histidine substitution was found to enhance the neutralization resistance of the spike glycoprotein in a chronical infection patient.[24] |
Summary
The constantly shifting epidemiology of coronavirus disease 2019 (COVID-19) ever since its initial outbreak has been a result of the continuous evolution of its etiological agent, SARS-CoV-2, from which numerous variants have been generated. Even within the first 2 years of this pandemic, the World Health Organization (WHO) has already announced 4 variants of concern (VOC), which are the previously circulating alpha (B.1.1.7), beta (B.1.351), gamma (P.1), and delta (B.1.617.2) strains, and many other variants of interest (VOI). The successive emergence of new SARS-CoV-2 variants has brought along many novel mutations, most of which continually refine and improve the fitness of the virus. For instance, these functionally advantageous mutations include the N501Y of alpha and L452R and E484Q of the B.1.617 lineage, which are capable of enhancing the ACE2-binding affinity of the spike glycoprotein.[25]
The latest SARS-CoV-2 lineage to be designated the status of VOC would be omicron (B.1.1.529) which first originated from South Africa.[26] This particular lineage alone has undergone substantial evolution over the course of its global dominance, spreading across the world like wildfire while simultaneously producing a diverse soup of dissimilar subvariants.[27] The first of its kind would be the BA.1 strain first appeared in November 2022. The supremacy of BA.1, however, would not last long, forasmuch as the emergence of the more fit BA.2 strain in December 2022 would eventually outcompete its antecedent.[28] Few months later, between March and July 2022, the successive emergences of BA.2.12.1, BA.4 and BA.5, and BA.2.75 would once again garnered the attention of the WHO and multiple countries. For one, the BA.2.12.1, BA.4, and BA.5 subvariants were found to possess enhanced antibody evasion capabilities and transmissibility when compared to the formerly active BA.2 strain,[27][29][30] allowing them to become dominant in the US and the UK.[31][32]BA.2.75, on the other hand, was the dominant variant in India, which habors higher hACE2-binding affinity than the BA.4 and BA.5 subvariants.[33][34] The complex interactions between these omicron sublineages prompted the creations of even more novel strains, including the recombinant XBB subvariant derived from BA.2.10.1 and BA.2.75 in August 2022, and the BQ.1 subvariant derived from BA.5 in October 2022. Like their predecessors, XBB swiftly rose to prominence upon its emergence, which was then succeeded by BQ.1 up till the end of 2022.[35][36] In fact, BQ.1.1, a descendent of BQ.1, was found to be the culprit behind 36.3% of the total US reported COVID-19 cases in December 2022.[37]
Recent emerging variant (EV) data retrieved from GISAID, as of 17 January 2023, has revealed that the top 4 most rapidly spreading lineages are the BA.1.1.22, CH.1.1, XBB.1.5, and BQ.1.1 variants, among which XBB.1.5 has been found to be especially prevalent in the US, making up of more than 40% of its sequence coverage in early January 2023.[37] The identified leading mutations are listed as follows:

References
- ↑ Jackson, C. B., Farzan, M., Chen, B. & Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat Rev Mol Cell Biol 23, 3–20 (2021).
- ↑ Weisblum, Y. et al. Escape from neutralizing antibodies by SARS-CoV-2 spike protein variants. eLife 9, (2020).
- ↑ Westendorf, K. et al. LY-CoV1404 (bebtelovimab) potently neutralizes SARS-CoV-2 variants. Cell Rep 39, 110812 (2022).
- ↑ 4.0 4.1 4.2 4.3 McCallum, M. et al. N-Terminal Domain Antigenic Mapping Reveals a Site of Vulnerability for SARS-CoV-2. Cell 184, 2332-2347 (2021).
- ↑ 5.0 5.1 Harvey, W. T. et al. SARS-CoV-2 variants, Spike mutations and immune escape. Nat Rev Microbiol 19, 409–424 (2021).
- ↑ Bugembe, D. L. et al. Emergence and spread of a SARS-COV-2 lineage a variant (A.23.1) with altered Spike Protein in Uganda. Nat Microbiol 6, 1094–1101 (2021).
- ↑ Oliva, R., Shaikh, A. R., Petta, A., Vangone, A. & Cavallo, L. D936Y and other mutations in the fusion core of the SARS-CoV-2 spike protein heptad repeat 1: Frequency, geographical distribution, and structural effect. Molecules 26, 1–13 (2021).
- ↑ Li, Q. et al. The Impact of Mutations in SARS-CoV-2 Spike on Viral Infectivity and Antigenicity. Cell 182, 1284-1294.e9 (2020).
- ↑ Donzelli, S. et al. Evidence of a SARS-CoV-2 double spike mutation D614G/S939F potentially affecting immune response of infected subjects. Comput Struct Biotechnol J 20, 733–744 (2022).
- ↑ Li, B. et al. Identification of Potential Binding Sites of Sialic Acids on the RBD Domain of SARS-CoV-2 Spike Protein. Front Chem. 9, 659764 (2021)
- ↑ 11.0 11.1 11.2 Zhou, L, et al. Predicting Spike Protein NTD Mutations of SARS-CoV-2 Causing Immune Evasion by Molecular Dynamics Simulations. Phys Chem Chem Phys 24, 3410–3419 (2022).
- ↑ Sun, X.-L. The role of cell surface sialic acids for SARS-CoV-2 infection. Glycobiology 31, 1245–1253 (2021).
- ↑ Buchanan, C. J. et al. Pathogen-sugar interactions revealed by Universal Saturation Transfer Analysis. Science 377, (2022).
- ↑ Suryadevara N. et al. An antibody targeting the N-terminal domain of SARS-CoV-2 disrupts the spike trimer. J Clin Invest 132, 159062 (2022).
- ↑ Wang, Q. et al. Key Mutations on Spike Protein Altering ACE2 Receptor Utilization and Potentially Expanding Host Range of Emerging SARS‐CoV‐2 Variants. J Med Virol. 95, 1-11 (2022).
- ↑ Gaebler, C. et al. Evolution of antibody immunity to SARS-CoV-2. Nature 591, 639–644 (2021).
- ↑ Wang, Q. et al. Resistance of SARS-CoV-2 omicron subvariant BA.4.6 to antibody neutralisation. Lancet Infect Dis 22, 1666–1668 (2022).
- ↑ Cong, Z. et al. Identification of SARS-CoV-2 spike mutations that attenuate monoclonal and serum antibody neutralization. Cell Host & Microbe 29, 1931-3128 (2021).
- ↑ 19.0 19.1 Greaney, A. et al. Comprehensive mapping of mutations in the SARS-CoV-2 receptor-binding domain that affect recognition by polyclonal human plasma antibodies. Cell Host Microbe 29, 463-476 (2021).
- ↑ Choi, Bina and Choudhary, Manish C. and Regan, James and Sparks, Jeffrey A. and Padera, Robert F. and Qiu, Xueting and Solomon, Isaac H. and Kuo, Hsiao-Hsuan and Boucau, Julie and Bowman, Kathryn and Adhikari, U. Das and Winkler, Marisa L. and Mueller, Al, J. Z. Persistence and Evolution of SARS-CoV-2 in an Immunocompromised Host. new engl J. Med. February, 2008–2009 (2020).
- ↑ Zhou, T. et al. Cryo-EM structures of SARS-CoV-2 spike without and with ACE2 reveal a pH-dependent switch to mediate endosomal positioning of receptor-binding domains. Cell Host Microbe 28, (2020).
- ↑ Henrich, S. et al. The crystal structure of the proprotein processing proteinase furin explains its stringent specificity. Nat Struct Mol Biol 10, 520–526 (2003).
- ↑ Tian, S. A 20 residues motif delineates the furin cleavage site and its physical properties may influence viral fusion. Biochem Insights 2, (2009).
- ↑ Kemp, S. A. et al. SARS-CoV-2 evolution during treatment of chronic infection. Nature 592, 277–282 (2021).
- ↑ Aggarwal, A. et al. Mechanistic Insights into the Effects of Key Mutations on SARS-CoV-2 RBD–ACE2 Binding. Phys Chem Chem Phys 23, 26451–26458 (2021)
- ↑ Karim, S. S. A. & Karim, Q. A. Omicron SARS-CoV-2 variant: A new chapter in the COVID-19 pandemic. Lancet 398, 2126–2128 (2021).
- ↑ 27.0 27.1 Tegally, H. et al. Emergence of SARS-CoV-2 omicron lineages BA.4 and BA.5 in South Africa. Nat Med 28, 1785–1790 (2022).
- ↑ Yamasoba, D. et al. Virological characteristics of the SARS-CoV-2 Omicron BA.2 spike. Cell 185, 2103-2115.e19 (2022).
- ↑ Cao, Y. et al. BA.2.12.1, BA.4 and BA.5 escape antibodies elicited by Omicron Infection. Nature 608, 593–602 (2022).
- ↑ Wang, Q. et al. Antibody evasion by SARS-CoV-2 omicron subvariants BA.2.12.1, BA.4 and BA.5. Nature 608, 603–608 (2022).
- ↑ Callaway, E. What Omicron’s BA.4 and BA.5 variants mean for the pandemic. Nature 606, 848–849 (2022).
- ↑ Del Rio, C. & Malani, P. N. COVID-19 in 2022 - The Beginning of the End or the End of the Beginning? JAMA 327, 2389–2390 (2022).
- ↑ Cao, Y. et al. Characterization of the enhanced infectivity and antibody evasion of Omicron BA.2.75. Cell Host Microbe 30, (2022).
- ↑ Shaheen, N. et al. Could the New BA.2.75 Sub-Variant Cause the Emergence of a Global Epidemic of COVID-19? A Scoping Review. Infect Drug Resist 15, 6317–6330 (2022).
- ↑ Wang, Q. et al. Alarming antibody evasion properties of rising SARS-CoV-2 BQ and XBB subvariants. Cell 186, (2023).
- ↑ European Centre for Disease Prevention and Control: Spread of the SARS-CoV-2 Omicron variant sub-lineage BQ.1 in the EU/EEA https://www.ecdc.europa.eu/sites/default/files/documents/Epi-update-BQ1.pdf (2022).
- ↑ 37.0 37.1 Highly immune evasive omicron XBB.1.5 variant is quickly becoming dominant in U.S. as it doubles weekly https://www.cnbc.com/2022/12/30/covid-news-omicron-xbbpoint1point5-is-highly-immune-evasive-and-binds-better-to-cells.html (2023).

Structure Testing